Síndrome del eutiroideo enfermo
Resumen breve
El hipotiroidismo se define como un estado de déficit crónico en la producción de hormonas tiroideas, lo que conduce a una disminución de la actividad metabólica tisular y al desarrollo de un conjunto de signos clínicos característicos. Desde el punto de vista etiopatogénico, el hipotiroidismo puede clasificarse como primario, cuando existe un fallo intrínseco de la glándula tiroides que impide la producción adecuada de tiroxina (T4) y trityodotironina (T3), o secundario, cuando la glándula…Índice de contenidos
Introducción
El hipotiroidismo se define como un estado de déficit crónico en la producción de hormonas tiroideas, lo que conduce a una disminución de la actividad metabólica tisular y al desarrollo de un conjunto de signos clínicos característicos. Desde el punto de vista etiopatogénico, el hipotiroidismo puede clasificarse como primario, cuando existe un fallo intrínseco de la glándula tiroides que impide la producción adecuada de tiroxina (T4) y trityodotironina (T3), o secundario, cuando la glándula tiroides es estructuralmente normal pero la secreción de hormona estimulante de la tiroides (TSH) se encuentra disminuida como consecuencia de una disfunción hipofisaria1-3. En consecuencia, la evaluación de la función tiroidea mediante la medición de hormonas tiroideas y TSH constituye un pilar fundamental para alcanzar un diagnóstico correcto y establecer una terapéutica adecuada. Sin embargo, la interpretación de estos parámetros no siempre resulta sencilla, ya que las concentraciones séricas de las hormonas tiroideas pueden verse alteradas por múltiples factores extratiroideos, lo que dificulta su correcta valoración clínica. Esta situación representa una de las principales causas de diagnóstico erróneo, particularmente de sobrediagnóstico de hipotiroidismo en pacientes que no presentan una enfermedad tiroidea primaria1-3. El síndrome del eutiroideo enfermo (SEE), también denominado síndrome del hipotiroideo enfermo o síndrome de enfermedad no tiroidea, se define como un conjunto de alteraciones en las concentraciones séricas de las hormonas tiroideas que se presentan en pacientes con enfermedades sistémicas agudas o crónicas, en ausencia de una patología de la glándula tiroides. En este contexto, el eje hipotálamo-hipófiso-tiroideo se encuentra funcionalmente alterado como consecuencia de la enfermedad subyacente, y no debido a un daño estructural o funcional intrínseco de la tiroides4-6. Desde el punto de vista fisiopatológico, el SEE se asocia a mecanismos complejos y multifactoriales que incluyen alteraciones en la conversión periférica de T4 a triyodotironina (T3), aumento de la producción de triyodotironina reversa (rT3), modificaciones en la unión de las hormonas tiroideas a proteínas transportadoras, cambios en la captación tisular y alteraciones en la secreción de TSH, influenciadas por citoquinas inflamatorias, ayuno prolongado y el estado metabólico del paciente4-6. En medicina veterinaria, este síndrome ha sido ampliamente documentado en perros con procesos inflamatorios sistémicos, infecciones graves, enfermedades endocrinas concurrentes, insuficiencia orgánica, neoplasias y estados críticos. Especialmente suele documentarse en situaciones crónicas de enfermedad. En estas situaciones, el SEE representa una de las principales causas de resultados de laboratorio falsamente compatibles con hipotiroidismo en animales que, en realidad, son eutiroideos, lo que puede conducir a decisiones terapéuticas inapropiadas1-6. Por lo tanto, para comprender adecuadamente por qué se produce el síndrome del eutiroideo enfermo y cómo evitar confundir a los pacientes afectados con verdaderos hipotiroideos, resulta imprescindible revisar en profundidad la bioquímica, la fisiología y la cinética de las hormonas tiroideas en el individuo normal, aspectos que serán desarrollados en los apartados siguientes.
Química estructural y síntesis de las hormonas tiroideas
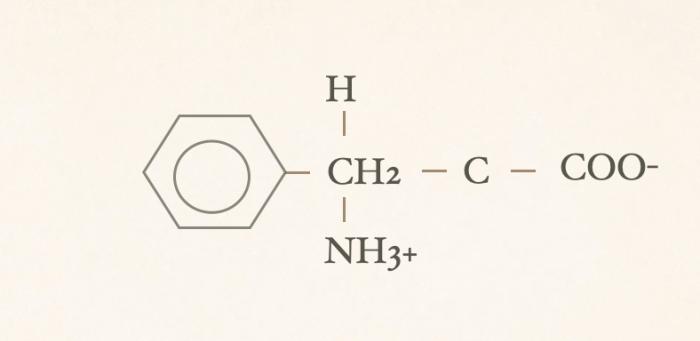
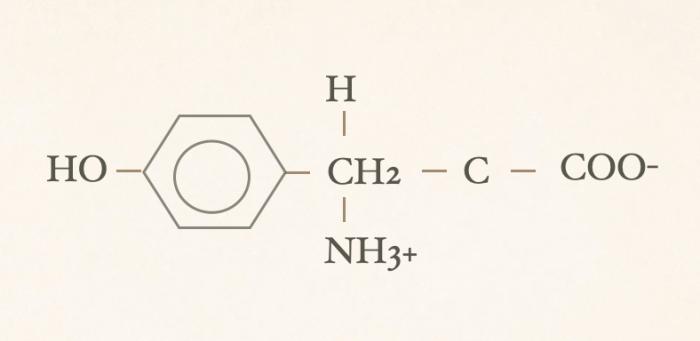
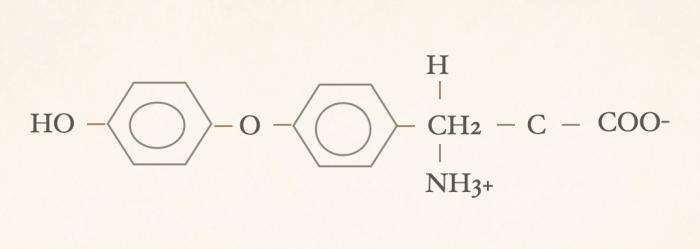
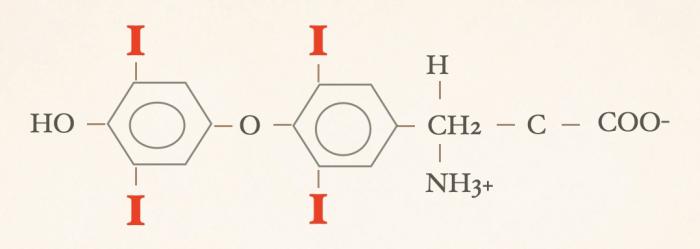
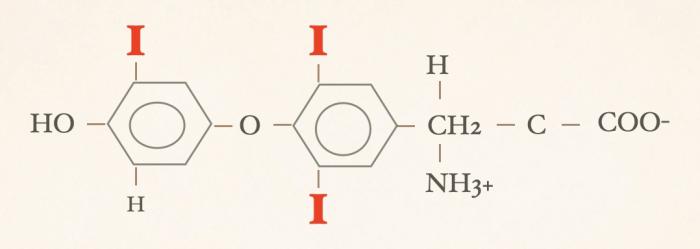
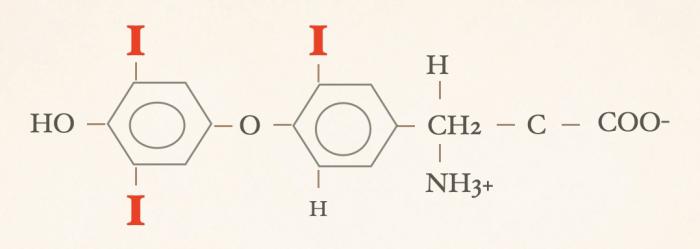
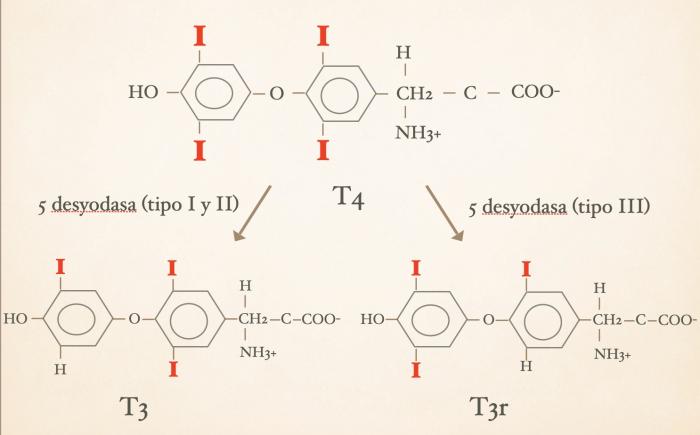
Las hormonas tiroideas son iodo-derivados orgánicos sintetizados y secretados por la glándula tiroides, cuya estructura básica se origina a partir del aminoácido tirosina (hidroxifenilalanina) (Figuras 1 y 2). Desde el punto de vista químico, pertenecen al grupo de las iodotironinas, y su síntesis ocurre en el interior del folículo tiroideo, específicamente en el coloide, como parte de una glicoproteína de gran tamaño denominada tiroglobulina, que actúa como molécula precursora y reservorio intraluminal de las hormonas tiroideas7,8. Estructuralmente, las hormonas tiroideas derivan de una molécula base denominada tironina (Figura 3), que representa el esqueleto químico común a todas ellas. La tironina corresponde a la condensación de dos anillos aromáticos derivados de residuos de tirosina (ver Figura 2) y no posee actividad biológica directa, ni se forma como molécula libre durante la síntesis fisiológica; su importancia radica en que constituye la base estructural sobre la cual se incorporan los átomos de yodo que dan origen a las iodotironinas biológicamente relevantes. Durante la síntesis hormonal, el yoduro captado por la célula folicular es transportado al lumen folicular y, bajo la acción de la tiroperoxidasa, se produce la organificación, es decir, la unión del yodo a residuos de tirosina presentes en la tiroglobulina. Posteriormente, el acoplamiento de estos residuos yodados permite la formación de las hormonas tiroideas. La incorporación de cuatro átomos de yodo a la estructura de la tironina da origen a la tetraiodotironina o tiroxina (T4) (Figura 4), mientras que la incorporación de tres átomos de yodo genera la triyodotironina (T3) (Figura 5). Ambas hormonas se sintetizan dentro del coloide tiroideo y permanecen almacenadas en la tiroglobulina hasta su liberación a la circulación sistémica7,8. En condiciones fisiológicas, la glándula tiroides produce principalmente T4 y, en menor proporción, T3, considerándose a la T4 el principal producto secretado y una prohormona desde el punto de vista funcional (la T4 carece de actividad biológica). También pueden generarse pequeñas cantidades de otras iodotironinas, como T2, las cuales carecen de relevancia biológica significativa en la regulación endocrina clásica7. La triyodotironina reversa (rT3) (Figura 6), si bien comparte la presencia de tres átomos de yodo con la T3, difiere en la posición de dichos átomos dentro de la molécula, lo que determina una actividad biológica mínima o nula. A diferencia de la tiroxina (T4), la triyodotironina reversa (rT3) no se sintetiza en la glándula tiroides, sino que se genera periféricamente a partir de la T4 mediante la desyodación del anillo interno de la molécula, proceso catalizado por enzimas iodotironina desyodinasas, principalmente del tipo 3 que conduce a la formación de un metabolito con actividad biológica mínima o nula, y adquiere especial relevancia en determinados estados fisiológicos y patológicos, en particular durante enfermedades sistémicas agudas o crónicas, como ocurre en el síndrome del eutiroideo enfermo, donde se observa un aumento relativo de rT3 como mecanismo adaptativo del metabolismo tiroideo4,5,7,9. Por su parte, la triyodotironina (T3) se sintetiza en parte en la glándula tiroides, aunque la mayor proporción de T3 circulante se produce en los tejidos periféricos (como hígado, riñón, sistema nervioso central, músculo y piel) a partir de la T4, mediante la desyodación del anillo externo catalizada por desyodinasas tipo 1 y 2 (Figura 7).Esta conversión periférica resulta crucial desde el punto de vista fisiológico, ya que la T3 representa la hormona tiroidea biológicamente activa, con una afinidad significativamente mayor por los receptores nucleares de hormonas tiroideas en comparación con la T4, siendo la principal responsable de los efectos metabólicos, tróficos y reguladores de la función celular atribuidos al eje tiroideo7,9. Todas estas desyodasas se encuentran principalmente en tejidos extratiroideos.
Coeficiente de partición lípido-agua: concepto general y aplicación a las hormonas tiroideas
El coeficiente de partición lípido-agua es un principio fisicoquímico que describe cómo una sustancia se distribuye entre dos fases que no se mezclan entre sí: una fase acuosa (agua) y una fase lipídica (grasa). Este concepto resulta esencial para comprender el comportamiento de numerosas moléculas biológicas, entre ellas las hormonas tiroideas. Desde el punto de vista conceptual, cuando una sustancia se encuentra simultáneamente en contacto con un medio acuoso y un medio lipídico, no se distribuye al azar, sino que lo hace siguiendo una proporción característica que depende de su estructura química. Esa proporción relativa entre ambas fases es lo que se denomina coeficiente de partición. De forma simplificada, puede pensarse que el coeficiente de partición indica qué fracción de una molécula “prefiere” el ambiente lipídico y qué fracción permanece en el ambiente acuoso.
¿Y las moléculas están fijas?Un aspecto fundamental (y muchas veces poco comprendido) del coeficiente de partición es que las moléculas en la fase acuosa y en la fase lipídica no están fijas, sino que se encuentran en un equilibrio dinámico.Por ejemplo, si una sustancia hipotética X presenta un coeficiente de partición tal que el 80 % de sus moléculas se encuentran en la fase lipídica y el 20 % en la fase acuosa, esto no significa que esas moléculas permanezcan inmóviles en cada compartimento. Por el contrario, existe un movimiento continuo de moléculas entre ambas fases, en ambas direcciones. En condiciones de equilibrio, la velocidad de paso desde la fase acuosa hacia la lipídica es igual a la velocidad en sentido contrario, por lo que la concentración en cada fase se mantiene constante. Este equilibrio es dinámico, no estático. Si el sistema se altera, el equilibrio tiende a restablecerse automáticamente. Si se retiran moléculas de la fase lipídica, disminuye su concentración, y esto provoca que moléculas desde la fase acuosa migren hacia la fase lipídica hasta recuperar la proporción 80–20. Si se retiran moléculas de la fase acuosa, ocurre el fenómeno inverso, con paso de moléculas desde la fase lipídica hacia la acuosa. Si se agregan moléculas a cualquiera de las dos fases, una parte de ellas se redistribuye hacia la otra fase hasta restablecer nuevamente la relación característica. Esto ocurre porque al modificar la concentración en una fase, se altera transitoriamente la velocidad de intercambio entre ambas, y el sistema responde desplazando moléculas hasta alcanzar nuevamente el equilibrio. En otras palabras, el coeficiente de partición no representa una distribución fija de moléculas, sino una relación de equilibrio estable que el sistema tiende a mantener constantemente.
Aplicación del coeficiente de partición a las hormonas tiroideas
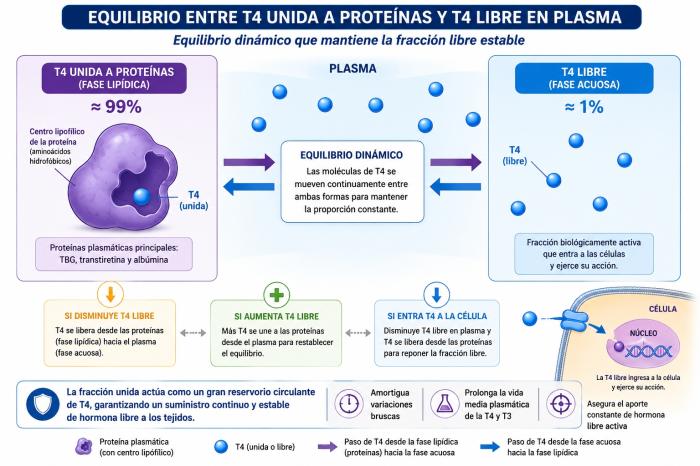
Las hormonas tiroideas, en particular la tiroxina (T4) y la triyodotironina (T3), constituyen un ejemplo muy ilustrativo de moléculas con un coeficiente de partición marcadamente elevado hacia la fase lipídica. Desde el punto de vista químico, ambas hormonas son altamente lipofílicas. Esto se debe a su estructura, rica en anillos aromáticos y átomos de yodo, lo que les confiere una fuerte afinidad por ambientes no acuosos. Como consecuencia, cuando son liberadas a la circulación, no permanecen principalmente disueltas en el plasma (que es un medio acuoso). En cambio, se unen de manera reversible a proteínas plasmáticas. Estas proteínas no son estructuras homogéneas: en su interior poseen regiones con características lipofílicas, formadas por aminoácidos hidrofóbicos. Estas regiones funcionan como pequeños “microambientes grasos” dentro de la proteína. De este modo, puede entenderse que las proteínas plasmáticas actúan como una especie de “fase lipídica transportadora” dentro del medio acuoso de la sangre. En condiciones fisiológicas aproximadamente 99 % de las hormonas tiroideas circulantes se encuentran unidas a proteínas plasmáticas, principalmente globulina fijadora de tiroxina (TBG), transtiretina y albúmina. Solo alrededor del 1 % (o incluso menos) se encuentra en forma libre, es decir, disuelta en la fase acuosa del plasma3,4.
Equilibrio entre hormona libre y hormona unida
Al igual que en el ejemplo teórico del 80-20, la distribución 99 % unida / 1 % libre no implica que la hormona unida y la libre sean compartimentos aislados. Por el contrario, ambas fracciones están en equilibrio dinámico permanente. Esto significa que si una molécula de T4 libre ingresa a una célula, disminuye momentáneamente la fracción libre en el plasma.De forma inmediata, una molécula de T4 previamente unida a proteínas se libera, pasando desde la fase “lipídica” (complejo proteína-hormona) a la fase acuosa, restaurando el equilibrio. Del mismo modo, si aumenta transitoriamente la concentración de hormona libre, una fracción se volverá a unir a proteínas plasmáticas. Este mecanismo asegura que la proporción entre hormona libre y hormona unida se mantenga notablemente estable, incluso frente a cambios en la captación tisular, metabolismo o excreción.
Significado fisiológico: un reservorio hormonal masivo
Desde esta perspectiva, la fracción de hormona tiroidea unida a proteínas plasmáticas no es una forma inactiva sin función, sino que cumple un rol fisiológico crucial. La T4 y la T3 unidas a proteínas actúan como un enorme reservorio circulante, capaz de amortiguar variaciones bruscas en la concentración de hormona libre, garantizar un suministro continuo de hormona a los tejidos y prolongar la vida media plasmática de las hormonas tiroideas (especialmente T4). De hecho, la hormona que efectivamente ingresa a las células y ejerce acción biológica es la fracción libre, pero esa fracción se mantiene gracias al constante aporte desde el compartimento unido, de acuerdo con el coeficiente de partición (Figura 8).
La T4 dentro del medio intracelular
El ingreso de T4 y T3 al interior celular no ocurre por simple difusión, sino mediante transportadores específicos de membrana, entre ellos MCT8, MCT10 y OATP1C1. La expresión de estos transportadores puede modificarse durante enfermedades sistémicas o inflamatorias, alterando la disponibilidad intracelular de hormonas tiroideas y contribuyendo al desarrollo del síndrome del eutiroideo enfermo4,10. La T4 que ingresa al medio intracelular posee escasa actividad biológica directa y, para ejercer plenamente sus efectos metabólicos, debe ser convertida a T3 mediante procesos de desyodación catalizados principalmente por las desyodasas tipo 1 (D1) y tipo 2 (D2). Estas enzimas remueven un átomo de yodo del anillo externo de la molécula de T4, generando T3, la forma hormonal metabólicamente más activa. Este proceso ocurre en numerosos tejidos periféricos, lo que hace que gran parte de la T3 no provenga directamente de la glándula tiroides, sino de la conversión periférica de T43,4,10. La T3 generada intracelularmente es transportada hacia el núcleo celular, donde se une a receptores nucleares específicos de hormonas tiroideas (TRα y TRβ). Estos receptores actúan como factores de transcripción capaces de modular la expresión de genes predeterminados en cada tipo celular, regulando procesos como metabolismo energético, termogénesis, síntesis proteica, diferenciación celular y actividad mitocondrial. La afinidad de los receptores nucleares por T3 es considerablemente mayor que por T4, razón por la cual la conversión intracelular constituye un paso esencial para la actividad biológica hormonal4,10. Una vez cumplidas sus funciones intracelulares, la T3 puede sufrir procesos de inactivación metabólica mediante desyodación adicional, especialmente por acción de la desyodasa tipo 3 (D3), que convierte T4 en T3 reversa (rT3) y T3 en metabolitos inactivos. Posteriormente, parte de la hormona retorna a la circulación sistémica, donde vuelve a establecerse el equilibrio dinámico entre las fracciones libres y las unidas a proteínas plasmáticas. Debido a que las hormonas tiroideas poseen una marcada lipofilia y más del 99 % circula unido a proteínas transportadoras, solo una pequeña fracción libre permanece biológicamente disponible para ingresar nuevamente a los tejidos3,4,9,10.
Bases fisiopatológicas del síndrome del eutiroideo enfermo
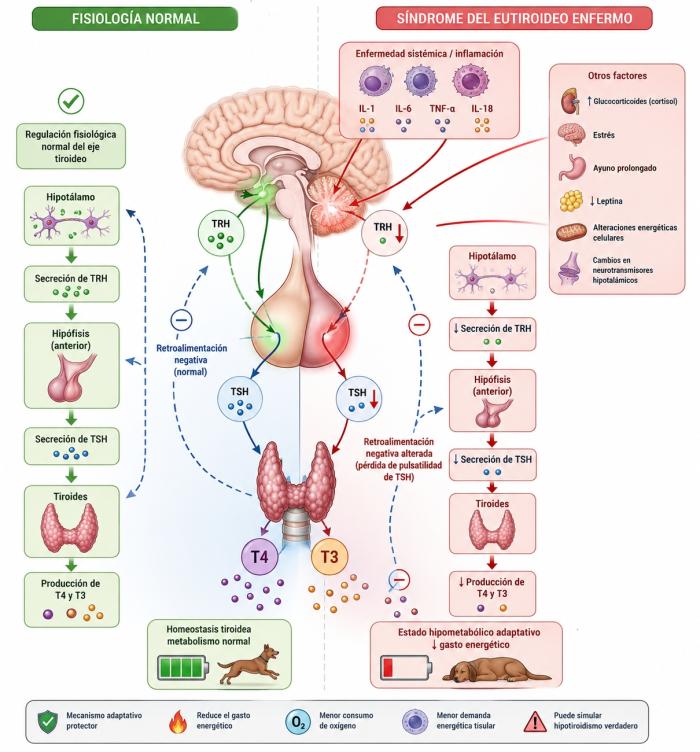
Se define como síndrome del eutiroideo enfermo al conjunto de alteraciones en las concentraciones circulantes de hormonas tiroideas que se producen como consecuencia de enfermedades no originadas en la glándula tiroides. En este cuadro, la glándula tiroides puede encontrarse estructural y funcionalmente normal, pero diversos mecanismos sistémicos asociados a inflamación, estrés metabólico o enfermedad crítica modifican la regulación del eje hipotálamo-hipófisis-tiroides y el metabolismo periférico de las hormonas tiroideas4,6,10. Debido a que gran parte del diagnóstico del hipotiroidismo se basa en la medición de las concentraciones séricas de T4, T3, T4 libre y TSH, las alteraciones inducidas por enfermedades extratiroideas pueden generar interpretaciones erróneas de los perfiles hormonales. Como consecuencia, el síndrome del eutiroideo enfermo constituye una de las principales causas de falsos diagnósticos de hipotiroidismo y, probablemente, una de las razones más frecuentes de administración innecesaria de levotiroxina en pacientes que no presentan una verdadera insuficiencia tiroidea primaria1-3,4,11. Las principales causas por las que puede ocurrir son:
- Disminución central del estímulo tiroideo: uno de los mecanismos más importantes implicados en el síndrome del eutiroideo enfermo es la alteración funcional del eje hipotálamo-hipófisis-tiroides (HHT). En este contexto, la disminución de las concentraciones circulantes de hormonas tiroideas no necesariamente refleja una falla primaria de la glándula tiroides, sino una reducción adaptativa del estímulo central ejercido sobre ella4,10. En condiciones fisiológicas normales, el hipotálamo secreta hormona liberadora de tirotropina (TRH), la cual estimula a las células tirotropas de la hipófisis para producir hormona estimulante de tiroides (TSH). A su vez, la TSH estimula la síntesis y liberación de T4 y T3 por la glándula tiroides. Este sistema funciona mediante mecanismos de retroalimentación negativa altamente regulados10. Durante enfermedades sistémicas, inflamatorias o estados críticos, múltiples señales metabólicas e inmunológicas alteran esta regulación central. Diversas citocinas proinflamatorias, particularmente interleucina-1 (IL-1), interleucina-6 (IL-6), factor de necrosis tumoral alfa (TNF-α) e interleucina-18 (IL-18), pueden actuar sobre el hipotálamo y la hipófisis disminuyendo la producción de TRH y modificando la secreción pulsátil de TSH4,10. Como consecuencia, el eje HHT entra en un estado funcional diferente, caracterizado por una menor estimulación tiroidea central. En muchos pacientes con síndrome del eutiroideo enfermo, la TSH puede encontrarse dentro del rango de referencia, discretamente disminuida o incluso levemente aumentada, pero frecuentemente pierde su patrón fisiológico normal de secreción pulsátil. Esto explica por qué la presencia de T4 disminuida junto con una TSH normal no necesariamente indica hipotiroidismo primario4,6,10. Estudios experimentales citados por Fliers y Boelen demostraron que procesos inflamatorios sistémicos inducen cambios simultáneos tanto en los componentes centrales como periféricos del eje tiroideo. La exposición a lipopolisacáridos bacterianos (LPS), por ejemplo, disminuye la expresión hipotalámica de TRH y altera la regulación neuroendocrina del eje HHT4,12. Además de las citocinas inflamatorias, otros factores asociados a enfermedad sistémica también participan en esta supresión central funcional, entre ellos glucocorticoides endógenos (síndrome se Cushing), aumento del cortisol asociado al estrés,, ayuno prolongado, disminución de leptina, alteraciones energéticas celulares y cambios en neurotransmisores hipotalámicos4,10. Las situaciones crónicas son las mas frecuentemente observadas aunque situaciones agudas son capaces tambien de inducir cambios significativos en las concentraciones de hormonas tiroideas. En enfermedades prolongadas o estados críticos severos, estas alteraciones centrales pueden profundizarse progresivamente, llevando a una disminución aún mayor de la secreción de TSH y contribuyendo al descenso de T4 y T3 circulantes4,8,12. Desde el punto de vista fisiopatológico, este fenómeno probablemente representa un mecanismo adaptativo destinado a reducir el gasto energético sistémico durante situaciones de estrés metabólico severo. La disminución del estímulo central tiroideo contribuiría así a generar un estado hipometabólico coordinado, con menor consumo de oxígeno y menor demanda energética tisular4,10. Uno de los aspectos clínicamente más relevantes de este mecanismo es que puede generar perfiles hormonales extremadamente similares a los observados en el hipotiroidismo verdadero, pese a que la glándula tiroides permanezca funcionalmente intacta. Esto constituye una de las principales bases fisiopatológicas de los falsos diagnósticos de hipotiroidismo en pacientes con enfermedades sistémicas1-3,6,11.
- Alteraciones en desyodasas con redirección del metabolismo hormonal de T4 - T3 hacia T4 - rT3: uno de los mecanismos fisiopatológicos centrales del síndrome del eutiroideo enfermo es la alteración del metabolismo periférico de las hormonas tiroideas, particularmente de la actividad de las enzimas desyodasas. Estas enzimas regulan la activación e inactivación de las hormonas tiroideas y determinan, en gran medida, la cantidad de T3 biológicamente activa disponible para los tejidos4,9,10. En condiciones fisiológicas normales, la mayor parte de la T3 circulante no es secretada directamente por la glándula tiroides, sino que se origina en tejidos periféricos a partir de la desyodación de T4. Este proceso es realizado principalmente por las desyodasas tipo 1 (D1) y tipo 2 (D2), las cuales remueven un átomo de yodo del anillo externo de la T4, transformándola en T3, la forma hormonal metabólicamente más activa. (2,22). Sin embargo, existe una tercera enzima, la desyodasa tipo 3 (D3), cuya función es distinta. La D3 remueve yodo del anillo interno de la T4, generando T3 reversa (rT3), una molécula estructuralmente similar a T3 pero con actividad biológica mínima o prácticamente ausente sobre los receptores nucleares tiroideos4,9,10. En estado fisiológico normal predomina la actividad de D1 y D2, favoreciendo la producción de T3. En cambio, durante enfermedades sistémicas, inflamación o estados críticos, ocurre una profunda reprogramación del metabolismo hormonal que disminuye la actividad de D1 y D2 y aumenta la actividad de D3. Como consecuencia se disminuye la conversión periférica de T4 hacia T3 y se aumenta la conversión de T4 hacia rT34,9,10. A raíz de esto descienden las concentraciones circulantes de T3, disminuyen progresivamente las concentraciones de T4 y aumentan las concentraciones de T3 reversa4,9. Este fenómeno representa uno de los hallazgos bioquímicos más característicos del síndrome del eutiroideo enfermo. Desde el punto de vista fisiopatológico esto implica que la disminución de T3 observada en estos pacientes no se origina necesariamente en una falla de producción tiroidea, sino en una redirección metabólica periférica del destino de la T4. En otras palabras, la T4 deja progresivamente de convertirse en hormona metabólicamente activa y pasa a transformarse preferentemente en metabolitos inactivos4,9,10. En la medida que la enfermedad progresa hacia la cronicidad se disminuyen también los niveles de T4 circulantes. Diversas citocinas inflamatorias participan en esta regulación de las desyodasas. Interleucina-1, interleucina-6, TNF-α y otras señales inflamatorias pueden modificar la expresión y actividad de estas enzimas en múltiples tejidos4,10. Además, el ayuno prolongado, el estrés metabólico, la enfermedad crítica y la disminución del aporte energético celular también favorecen este cambio enzimático4,10.Desde una perspectiva adaptativa, esta modificación del metabolismo hormonal probablemente constituye un mecanismo destinado a reducir el gasto energético del organismo durante períodos de enfermedad severa(al igual que con la disminución central del eje tiroideo). La disminución de T3 intracelular reduce el metabolismo basal, la termogénesis, el consumo de oxígeno y múltiples procesos anabólicos celulares4,10. Un aspecto particularmente importante es que estos cambios pueden ocurrir aun cuando la glándula tiroides permanezca completamente funcional. Por ello, concentraciones bajas de T3 o T4 durante enfermedad sistémica no necesariamente reflejan hipotiroidismo verdadero, sino una modificación adaptativa del metabolismo periférico hormonal4,6,10. La elevación de la T3 reversa (T3r) adquiere aquí especial relevancia fisiopatológica. Debido a que el rT3 se produce a expensas de la T4 que normalmente sería convertida en T3 activa, su aumento constituye un marcador indirecto del desvío metabólico característico del síndrome del eutiroideo enfermo9. En conjunto, las alteraciones de las desyodasas representan uno de los mecanismos moleculares más importantes responsables de las modificaciones hormonales observadas en pacientes con enfermedades sistémicas no tiroideas.
- Alteraciones en proteínas transportadoras: otro de los mecanismos fundamentales implicados en el síndrome del eutiroideo enfermo es la alteración de las proteínas plasmáticas transportadoras de hormonas tiroideas y de la dinámica de unión hormonal en sangre. Debido a que más del 99 % de la T4 y T3 circula unida a proteínas plasmáticas, pequeñas modificaciones en este sistema pueden producir cambios significativos en las concentraciones hormonales medidas en laboratorio, aun cuando la glándula tiroides permanezca funcionalmente normal3,4,10. Las principales proteínas transportadoras de hormonas tiroideas son la globulina fijadora de tiroxina (TBG), la transtiretina y la albúmina4,9,10. Estas proteínas poseen regiones internas con características lipofílicas capaces de unirse reversiblemente a T4 y T3. Desde el punto de vista fisicoquímico, estos complejos proteína-hormona funcionan como una “fase lipídica circulante”, mientras que la pequeña fracción libre de hormona representa la fase acuosa plasmática. En condiciones normales, ambas fracciones permanecen en equilibrio dinámico constante. Cuando una molécula libre ingresa a una célula o es metabolizada, una fracción de hormona previamente unida se libera desde las proteínas plasmáticas para restablecer el equilibrio. Gracias a este mecanismo, la enorme masa hormonal unida a proteínas actúa como un gigantesco reservorio circulante3,4,10. Durante enfermedades sistémicas, múltiples alteraciones pueden modificar este equilibrio disminución en la síntesis de proteínas transportadoras,cambios en la afinidad de unión hormonal, desplazamiento competitivo por otras moléculas, modificaciones estructurales proteicas inducidas por inflamación, alteraciones hepáticas, cambios en albúmina plasmática y presencia de fármacos que interfieren con la unión hormonal4,6,7,10,13. Como consecuencia, puede disminuir la cantidad de hormona unida a proteínas y alterarse la distribución entre las fracciones libre y ligada. Un aspecto particularmente importante es que la concentración de T4 total depende en gran medida de la cantidad de hormona unida a proteínas plasmáticas. Por ello, alteraciones en las proteínas transportadoras pueden producir disminuciones marcadas de T4 total sin que inicialmente exista una reducción equivalente de la hormona libre biológicamente activa3,14-16. Esto explica por qué muchos pacientes con enfermedades sistémicas pueden presentar valores bajos de T4 total pese a no padecer un verdadero hipotiroidismo primario. Además, durante procesos inflamatorios severos, las proteínas transportadoras podrían disminuir su síntesis hepática. En paralelo, diversas moléculas circulantes, ácidos grasos libres, metabolitos y medicamentos pueden competir con las hormonas tiroideas por sus sitios de unión plasmáticos, alterando aún más el equilibrio hormonal4,7,10,13. Entre las drogas reportadas como modificadoras de este equilibrio figuran glucocorticoides, fenobarbital, sulfonamidas potenciadas y amiodaronaentre otras7,17. Desde el punto de vista conceptual, esto puede entenderse como una modificación del coeficiente de partición hormonal entre la fase unida y la fase libre. Si disminuye la capacidad de la “fase lipídica transportadora” para alojar hormonas tiroideas, el sistema redistribuye temporalmente las moléculas hasta alcanzar un nuevo equilibrio dinámico. Sin embargo, debido a que las hormonas libres son rápidamente captadas, metabolizadas o eliminadas, estas modificaciones pueden terminar acompañándose también de disminuciones progresivas de las concentraciones hormonales circulantes totales y libres4,6,10. Este mecanismo posee enorme relevancia clínica porque gran parte de las pruebas hormonales utilizadas en medicina veterinaria cuantifican T4 total. En consecuencia, alteraciones de las proteínas transportadoras constituyen una de las principales causas de perfiles hormonales compatibles con hipotiroidismo en pacientes que en realidad presentan enfermedades extratiroideas2-4,6,11.
- Disminución del ingreso celular de hormonas tiroideas: durante muchos años se consideró que las hormonas tiroideas atravesaban las membranas celulares principalmente por difusión pasiva debido a su naturaleza lipofílica. Sin embargo, actualmente se sabe que el ingreso de T4 y T3 al interior celular depende en gran medida de transportadores específicos presentes en la membrana plasmática4,10. Este concepto posee gran relevancia en el síndrome del eutiroideo enfermo, ya que la actividad biológica de las hormonas tiroideas no depende únicamente de sus concentraciones circulantes, sino también de la capacidad de los tejidos para captar dichas hormonas desde el medio extracelular4,10. Entre los principales transportadores identificados para hormonas tiroideas se encuentran: MCT8 (monocarboxylate transporter 8), MCT10, OATP1C1 y otros transportadores tisulares especializados4,10. Estos sistemas regulan el paso de T4 y T3 hacia el interior celular, permitiendo posteriormente su conversión intracelular mediante desyodasas y su posterior interacción con receptores nucleares4,10. Durante enfermedades sistémicas, inflamación o estados críticos, la expresión y actividad funcional de estos transportadores puede alterarse significativamente. Diversas citocinas inflamatorias y señales asociadas al estrés metabólico modifican la captación tisular de hormonas tiroideas, disminuyendo su disponibilidad intracelular4,10. Como consecuencia, algunos tejidos pueden experimentar una reducción funcional de la actividad tiroidea intracelular aun cuando las concentraciones plasmáticas hormonales no se encuentren marcadamente disminuidas. Desde el punto de vista fisiopatológico, esto implica que durante el síndrome del eutiroideo enfermo puede desarrollarse una forma de “hipotiroidismo tisular relativo”, originado no necesariamente por falta de hormona circulante, sino por una menor capacidad celular para incorporarla4,10. Además, distintos tejidos expresan diferentes tipos y cantidades de transportadores, por lo que las alteraciones del transporte hormonal pueden variar considerablemente entre órganos. Esto contribuye a la heterogeneidad metabólica observada durante enfermedades sistémicas severas4,10.
- Resistencia tisular relativa a las hormonas tiroideas: la actividad biológica de las hormonas tiroideas no depende únicamente de su ingreso al interior celular. Para ejercer sus efectos metabólicos, la T3 debe interactuar con receptores nucleares específicos capaces de regular la expresión génica en múltiples tejidos. Por este motivo, alteraciones en los receptores tiroideos o en sus mecanismos de señalización intracelular también pueden contribuir significativamente a la fisiopatología del síndrome del eutiroideo enfermo4,10. Los principales receptores de hormonas tiroideas pertenecen a la familia de receptores nucleares TR (thyroid hormone receptors), fundamentalmente TRα y TRβ. Estos receptores funcionan como factores de transcripción que, una vez unidos a T3, modulan la expresión de genes específicos involucrados en el metabolismo energético, el consumo de oxígeno, la función mitocondrial, la síntesis proteica, la termogénesis, el crecimiento, la diferenciación celular y numerosas funciones metabólicas tisulares10. La afinidad de estos receptores por T3 es considerablemente mayor que por T4, razón por la cual gran parte de la actividad hormonal depende de la conversión intracelular de T4 en T3 mediante las desyodasas4,10. Durante enfermedades sistémicas, inflamación o estados críticos, diversos mecanismos pueden alterar la respuesta tisular a las hormonas tiroideas aun cuando existan concentraciones circulantes detectables de hormona. Este fenómeno ha sido interpretado como una forma de “resistencia tisular relativa” a las hormonas tiroideas4,10. Diversas citocinas inflamatorias, particularmente IL-1, IL-6 y TNF-α, pueden modificar la expresión y actividad funcional de los receptores tiroideos y de múltiples proteínas involucradas en la señalización nuclear hormonal4,10. Además, durante inflamación sistémica se producen alteraciones en coactivadores transcripcionales, correpresores nucleares, mecanismos de unión receptor-ADN, función mitocondrial y vías intracelulares dependientes de energía10. Como consecuencia, puede disminuir la respuesta biológica celular a las hormonas tiroideas aun sin existir una deficiencia hormonal absoluta. Desde el punto de vista fisiopatológico, esto implica que durante el síndrome del eutiroideo enfermo no solo puede reducirse la cantidad de hormona disponible para los tejidos, sino también la capacidad de las células para responder adecuadamente a ella4,10.Este concepto resulta particularmente importante porque ayuda a explicar por qué algunos pacientes presentan manifestaciones metabólicas compatibles con un estado hipotiroideo pese a mantener concentraciones hormonales circulantes relativamente conservadas.
- Modificaciones inducidas por fármacos sobre el eje y metabolismo tiroideo: diversos fármacos pueden producir alteraciones significativas en las concentraciones circulantes de hormonas tiroideas y en la regulación del eje hipotálamo-hipófisis-tiroides, aun en ausencia de una enfermedad tiroidea primaria. Estas modificaciones poseen gran importancia clínica, ya que constituyen una de las principales causas de perfiles hormonales compatibles con hipotiroidismo en pacientes que en realidad presentan efectos farmacológicos reversibles sobre la homeostasis tiroidea3,7,17. Un aspecto particularmente relevante es que la mayoría de los medicamentos no actúa mediante un único mecanismo aislado. Por el contrario, muchas drogas interfieren simultáneamente con la secreción de TSH, la actividad de desyodasas, el metabolismo hepático hormonal, la unión a proteínas transportadoras, la depuración hormonal y la disponibilidad periférica de T4 y T37,17. Por este motivo, las alteraciones inducidas por fármacos suelen ser multifactoriales y pueden generar perfiles hormonales complejos.
- Glucocorticoides: los glucocorticoides constituyen uno de los grupos farmacológicos más frecuentemente asociados a alteraciones tiroideas. Tanto el cortisol endógeno asociado al estrés como la administración exógena de corticoides pueden disminuir las concentraciones séricas de T4 total y, en menor medida, de T33,7,17. Entre los mecanismos involucrados se incluyen disminución de la secreción de TSH, alteración de la conversión periférica de T4 hacia T3, modificaciones en proteínas transportadoras y aumento del metabolismo hormonal periférico7,17. Daminet y colaboradores demostraron que incluso dosis antiinflamatorias de prednisona pueden producir descensos significativos de T4 en perros hipotiroideos bajo tratamiento, modificando la interpretación de las pruebas hormonales17.
- Anticonvulsivantes: el fenobarbital y otros anticonvulsivantes también pueden producir disminuciones importantes de T4 total y T4 libre3,7. El principal mecanismo involucrado parece ser la inducción enzimática hepática, con aumento del metabolismo y depuración de hormonas tiroideas. Además, algunos anticonvulsivantes pueden modificar indirectamente la unión hormonal a proteínas plasmáticas y alterar el metabolismo periférico de las hormonas tiroideas7. Estas alteraciones pueden ser suficientemente marcadas como para generar perfiles bioquímicos compatibles con hipotiroidismo pese a que la glándula tiroides permanezca funcionalmente normal3,7.
- Sulfonamidas potenciadas: las sulfonamidas potenciadas poseen un efecto particularmente importante sobre la función tiroidea y representan uno de los fármacos más asociados a alteraciones hormonales significativas en perros3,7. Estos medicamentos pueden interferir directamente con la síntesis hormonal tiroidea, alterar la organificación del yodo y producir disminuciones marcadas de T4 y T3 circulantes7. En algunos pacientes, los cambios inducidos por sulfonamidas pueden incluso acompañarse de elevaciones de TSH, generando perfiles hormonales extremadamente similares a los observados en el hipotiroidismo primario verdadero3,7. En algunos pacientes, las alteraciones inducidas por sulfonamidas pueden acompañarse de un incremento en las concentraciones séricas de TSH, produciendo un cuadro de hipotiroidismo secundario a dishormonogénesis tiroidea adquirida. Este efecto suele ser reversible tras la suspensión del fármaco7.
- Toceranib y otros fármacos oncológicos: algunos agentes antineoplásicos, particularmente inhibidores de tirosina quinasa como toceranib, también han sido asociados a alteraciones hormonales tiroideas. Harper y colaboradores documentaron modificaciones en las concentraciones de hormonas tiroideas en perros tratados con toceranib, sugiriendo posibles efectos sobre el metabolismo hormonal periférico y el eje tiroideo12.
- Amiodarona y alteraciones del metabolismo periférico: la amiodarona posee una compleja interacción con el metabolismo tiroideo debido a su elevado contenido de yodo y a su capacidad para modificar la actividad de desyodasas periféricas. En humanos, se ha asociado particularmente con alteraciones en la conversión de T4 hacia T3 y aumento de T3r (T3 Reversa)9. En perros, la administración de amiodarona se ha asociado con la aparición de hipotiroidismo iatrogénico reversible, el cual generalmente remite tras la suspensión del tratamiento.
Desde el punto de vista fisiopatológico, las alteraciones farmacológicas demuestran que las concentraciones hormonales tiroideas pueden modificarse significativamente sin que exista una enfermedad primaria de la glándula tiroides. Esto refuerza el concepto de que las pruebas hormonales deben interpretarse siempre en función del contexto clínico completo del paciente1-3,11. Además, muchos de estos cambios farmacológicos reproducen mecanismos similares a los observados en el síndrome del eutiroideo enfermo, incluyendo disminución de TSH, alteración de desyodasas, cambios en proteínas transportadoras y modificaciones del metabolismo periférico hormonal7,17. Por este motivo, el conocimiento detallado de las interacciones farmacológicas sobre el sistema tiroideo resulta esencial para evitar diagnósticos erróneos de hipotiroidismo y tratamientos innecesarios con levotiroxina1-3,7,11.
- Reprogramación metabólica durante enfermedad sistémica: actualmente, el síndrome del eutiroideo enfermo no se interpreta únicamente como una alteración pasiva de las hormonas tiroideas secundaria a enfermedad sistémica, sino como una compleja respuesta adaptativa del organismo frente al estrés metabólico, la inflamación y la enfermedad crítica4,10. Desde esta perspectiva, muchas de las modificaciones observadas en el eje tiroideo y en el metabolismo hormonal representarían mecanismos coordinados destinados a disminuir el gasto energético celular y favorecer la supervivencia durante períodos de agresión sistémica severa4,10. Las hormonas tiroideas cumplen un rol central en la regulación del metabolismo basal. Entre sus múltiples efectos fisiológicos se incluyen aumento del consumo de oxígeno, estimulación de la actividad mitocondrial, incremento de la termogénesis, regulación de la síntesis y degradación proteica, y aumento del gasto energético tisular10. Por este motivo, una reducción funcional de la actividad tiroidea puede disminuir considerablemente las demandas metabólicas del organismo. Durante enfermedades sistémicas severas, inflamación, sepsis, trauma, ayuno prolongado o estados críticos, el organismo activa múltiples mecanismos orientados a conservar energía. Entre ellos se incluyen disminución de TRH y TSH,reducción de la conversión periférica de T4 hacia T3, aumento de reverse T3, disminución del transporte intracelular hormonal y alteraciones en la respuesta nuclear a hormonas tiroideas4,10. Todos estos mecanismos convergen funcionalmente en un mismo resultado: la disminución global de la actividad metabólica tisular. Desde un punto de vista fisiopatológico moderno, el síndrome del eutiroideo enfermo puede entenderse entonces como una forma de “reprogramación metabólica coordinada”, en la cual el organismo reduce transitoriamente múltiples procesos consumidores de energía con el objetivo de priorizar funciones vitales esenciales4,10. Esta respuesta probablemente posee un importante valor adaptativo durante fases agudas de enfermedad. La reducción del metabolismo basal puede disminuir el consumo energético, reducir requerimientos de oxígeno, limitar el catabolismo tisular, y mejorar la redistribución de recursos metabólicos hacia órganos críticos y mecanismos inmunológicos4,10. Diversos trabajos experimentales y clínicos han demostrado que las disminuciones de T3 observadas durante enfermedad sistémica se correlacionan frecuentemente con la gravedad del cuadro clínico5,6,13,18. En perros críticamente enfermos, las alteraciones hormonales más marcadas suelen observarse en pacientes con mayor compromiso sistémico, lo que refuerza la idea de que estos cambios forman parte de una respuesta metabólica global del organismo frente al estrés severo5,6,18. Sin embargo, aún existe debate acerca de si esta respuesta permanece completamente adaptativa durante enfermedades prolongadas o extremadamente graves. Algunos autores sugieren que, en estados críticos persistentes, la supresión mantenida del eje tiroideo podría eventualmente contribuir al deterioro metabólico y a la disfunción orgánica4,10. Este concepto posee gran relevancia clínica, ya que implica que las alteraciones hormonales observadas durante enfermedad sistémica no necesariamente representan una deficiencia primaria de hormonas tiroideas que deba corregirse farmacológicamente. Por el contrario, en muchos casos podrían constituir parte de una estrategia fisiológica adaptativa compleja desarrollada para favorecer la supervivencia durante períodos de estrés metabólico severo4,10 (Figura 9).
¿Qué hormonas tiroideas se modifican en el síndrome del eutiroideo enfermo?
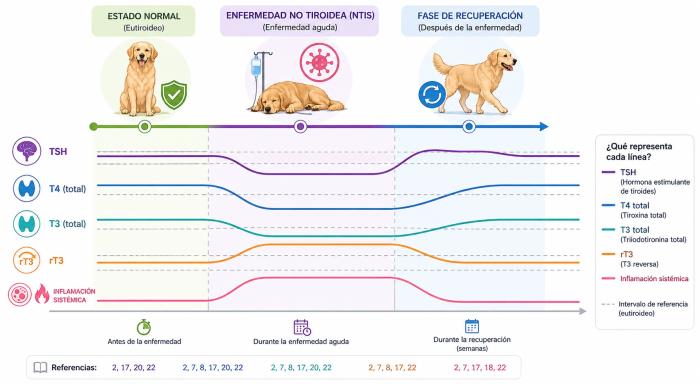
Las alteraciones hormonales observadas en el síndrome del eutiroideo enfermo no ocurren de manera simultánea ni con la misma intensidad. A medida que la enfermedad sistémica progresa, diferentes mecanismos fisiopatológicos afectan secuencialmente el eje hipotálamo-hipófisis-tiroides, el metabolismo periférico hormonal y la disponibilidad tisular de hormonas tiroideas. Como consecuencia, las distintas hormonas y metabolitos tiroideos presentan patrones de alteración relativamente característicos4,6,9,10.
- T3 (Triyodotironina): la T3 suele ser la primera hormona en disminuir y constituye una de las alteraciones bioquímicas más tempranas y constantes del síndrome del eutiroideo enfermo4,6,9,10. La principal causa de esta disminución es la reducción de la conversión periférica de T4 hacia T3 secundaria a disminución de la actividad de las desyodasas tipo 1 (D1) y tipo 2 (D2), junto con un aumento relativo de la actividad de la desyodasa tipo 3 (D3)4,9,10. Debido a que gran parte de la T3 circulante se origina en tejidos periféricos y no directamente en la glándula tiroides, las alteraciones en las desyodasas producen rápidamente un descenso de sus concentraciones séricas4,10. En pacientes críticamente enfermos, la magnitud de la disminución de T3 suele correlacionarse con la severidad del cuadro sistémico y con el pronóstico clínico5,6,13,18.
- T3 reversa (rT3): la T3 reversa (rT3) suele aumentar de manera relativamente sincrónica con la disminución de T3 y representa uno de los hallazgos metabólicos más característicos del síndrome del eutiroideo enfermo4,9,10. La rT3 se origina principalmente por desyodación interna de T4 mediada por la desyodasa tipo 3 (D3). Durante enfermedad sistémica, disminuye la conversión de T4 hacia T3 y aumenta su conversión hacia rT3, produciendo un “desvío metabólico” desde hormonas biológicamente activas hacia metabolitos inactivos4,9,15. Además, la disminución de la actividad de D1 reduce la depuración periférica de rT3, favoreciendo aún más su acumulación sérica9,15. Aunque la rT3 constituye un marcador fisiopatológico muy interesante del síndrome del eutiroideo enfermo, su utilización clínica rutinaria continúa siendo limitada tanto en medicina humana como veterinaria9.
- T4 total (TT4): la T4 total suele disminuir más tardíamente y, en general, sus descensos se asocian a enfermedades sistémicas más severas, prolongadas o críticas4,6,10. Su disminución puede originarse por múltiples mecanismos simultáneos, entre ellos disminución del estímulo hipotalámico-hipofisario, reducción de la secreción tiroidea, alteraciones en proteínas transportadoras, aumento del metabolismo periférico y modificaciones en la conversión hormonal4,6,10. Debido a que más del 99 % de la T4 circula unida a proteínas plasmáticas, pequeñas alteraciones en la unión hormonal pueden producir disminuciones importantes de TT4 sin que exista inicialmente una caída proporcional de la hormona libre biológicamente activa3,15,16. Por este motivo, concentraciones bajas de TT4 constituyen uno de los principales factores responsables de falsos diagnósticos de hipotiroidismo en pacientes con enfermedades extratiroideas1-3,6,11.
- T4 libre (fT4): la T4 libre suele preservarse durante más tiempo y generalmente se altera en fases más avanzadas o severas del síndrome del eutiroideo enfermo6,15,16. Sin embargo, la confiabilidad diagnóstica de la fT4 depende enormemente de la técnica utilizada para su medición. Cuando se determina mediante diálisis de equilibrio (equilibrium dialysis), la fT4 resulta considerablemente menos afectada por enfermedades extratiroideas y por alteraciones en proteínas transportadoras plasmáticas15,16,18. Por el contrario, diversos inmunoensayos analógicos pueden subestimar la concentración real de fT4 y mostrar disminuciones paralelas a la TT4 debido a interferencias metodológicas y alteraciones en el equilibrio proteína-hormona15,16. Por este motivo, la medición de fT4 mediante diálisis de equilibrio es considerada una de las pruebas más útiles para diferenciar hipotiroidismo verdadero de síndrome del eutiroideo enfermo en perros3,6,15,16.
- TSH canina (cTSH): La TSH canina suele ser la hormona menos afectada y la última en modificarse durante el síndrome del eutiroideo enfermo3,4,6,10. A diferencia del hipotiroidismo primario verdadero, en el síndrome del eutiroideo enfermo generalmente no se observa una elevación marcada de TSH pese a la disminución de T4 y T3. Esto se debe a alteraciones funcionales del eje hipotálamo-hipófisis-tiroides, con disminución de TRH hipotalámica y pérdida parcial de la secreción fisiológica pulsátil de TSH4,6,10. En algunos pacientes, la TSH puede encontrarsedentro del rango de referencia, discretamente disminuida, o levemente elevada3,6. En cuadros extremadamente severos o prolongados puede observarse una supresión más marcada de TSH4,10. Durante la fase de recuperación del síndrome del eutiroideo enfermo, es relativamente frecuente observar elevaciones transitorias de TSH, probablemente reflejando la recuperación progresiva de la funcionalidad normal del eje hipotálamo-hipófisis-tiroides y el restablecimiento de la estimulación tiroidea fisiológica4. Es importante recordad en este punto que una THS elevada no necesariamente confirma hipotiroidismo real, ya que ha individuos normotiroideos con niveles de TSH anormalmente elevados (Figura 10).
¿Entonces cómo se diferencia un hipotiroidismo verdadero de un síndrome del eutiroideo enfermo?
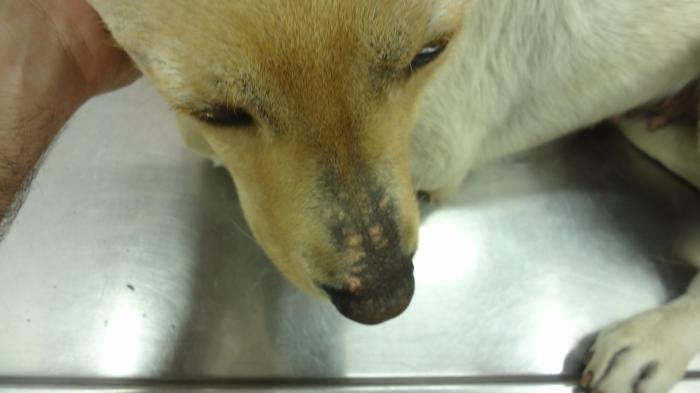
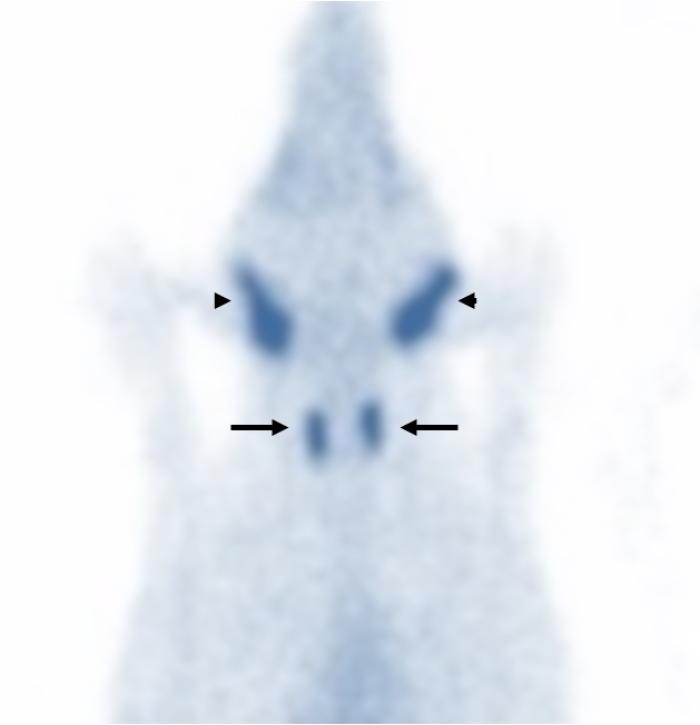
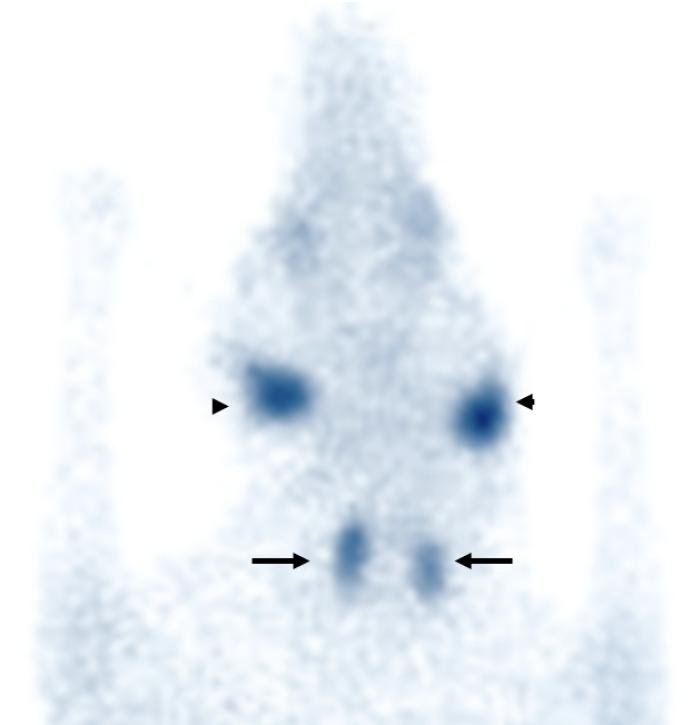
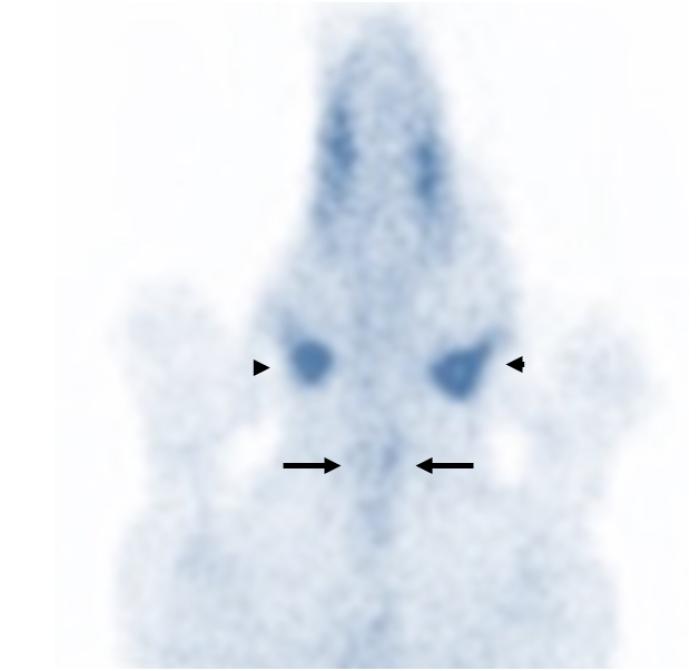
La diferenciación entre un hipotiroidismo verdadero y un síndrome del eutiroideo enfermo constituye uno de los mayores desafíos diagnósticos de la endocrinología veterinaria. Esto se debe a que ambos procesos pueden producir alteraciones hormonales muy similares, particularmente disminuciones de T4 total, T3 e incluso T4 libre o alteraciones en TSHc1-3,6,11. Por este motivo, el diagnóstico de hipotiroidismo nunca debe sustentarse exclusivamente en la medición aislada de hormonas tiroideas. La interpretación correcta requiere integrar la historia clínica, los hallazgos físicos, las alteraciones dermatológicas y en otros sistemas frecuentemente afectados como el ojo o el sistema nervioso,los cambios bioquímicos compatibles, las pruebas hormonales, y la evolución clínica del paciente1-3,11. Los perros con hipotiroidismo verdadero suelen presentar manifestaciones sistémicas compatibles con disminución del metabolismo basal, entre ellas letargia, intolerancia al ejercicio, tendencia al aumento de peso, intolerancia al frío, disminución de la actividad, bradicardia, y alteraciones reproductivas1-3,8,11. Desde el punto de vista dermatológico, las alteraciones cutáneas representan uno de los motivos de consulta más frecuentes y pueden incluir pelaje seco o de mala calidad(Figura 11), alopecia del puente nasal (hallazgo muy específico para Hipotiroidismo) (Figuras 12 y 13), alopecia caudal (“cola de rata”) (Figuras 12 y 14), hiperpigmentación, hiperqueratosis, seborrea y predisposición a infecciones cutáneas secundarias1-3,8,11. La alopecia bilateral simétrica extensa y las otitis recurrentes severas son hallazgos frecuentemente sobredimensionados en relación con el hipotiroidismo canino y constituyen una causa importante de errores diagnósticos en la práctica clínica. Alopecia completa simétrica y bilateral se observa en otras endocrinopatías pero no en hipotiroidismo, donde el pelaje de mala calidad es el hallazgo cutáneo más característico en relación con Hipotiroidismo. En realidad, muchos pacientes con grandes cuadros de alopecia generalizada u otitis recidivantes crónicas presentan otras enfermedades dermatológicas o sistémicas que alteran secundariamente el perfil hormonal tiroideo y generan un síndrome del eutiroideo enfermo1-3,6. Asimismo, numerosas alteraciones cutáneas compatibles con hipotiroidismo pueden observarse también en pacientes con procesos inflamatorios crónicos, endocrinopatías diferentes, enfermedades sistémicas severas o dermatosis primarias no endocrinas1-3,6. Por este motivo, la presencia de lesiones dermatológicas junto con alteraciones hormonales tiroideas no confirma por sí sola un hipotiroidismo verdadero y siempre debe interpretarse dentro del contexto clínico completo del paciente1-3,6,11. Por ello, la simple presencia de TT4 disminuida asociada a signos dermatológicos no confirma por sí sola un hipotiroidismo verdadero. De hecho, uno de los errores diagnósticos más frecuentes consiste en interpretar automáticamente una T4 baja como evidencia definitiva de insuficiencia tiroidea1-3,11. La evaluación hormonal debe realizarse siempre considerando el contexto clínico completo del paciente. En general, valores marcadamente bajos de TT4 incrementan la sospecha diagnóstica de hipotiroidismo, mientras que concentraciones normales hacen que esta enfermedad sea poco probable1,3. Sin embargo, debido a la influencia que pueden ejercer las enfermedades extratiroideas y diversos medicamentos sobre la TT4, con frecuencia es necesario complementar la evaluación con pruebas dinámica e imágenes morfofuncionales.Entre estas, la estimulación con TSH recombinante humana, la prueba de estimulación con TRH y la gammagrafía han sido consideradas métodos de referencia para la valoración funcional tiroidea. La medición de T4 libre mediante diálisis de equilibrio presenta una alta especificidad diagnóstica; no obstante, incluso esta prueba puede verse afectada en pacientes con enfermedades sistémicas severas. Por otra parte, los anticuerpos antitiroglobulina pueden detectarse en perros con tiroiditis sin hipotiroidismo clínico, así como pueden ser negativos en animales con hipotiroidismo crónico. En este contexto, dichos anticuerpos se consideran principalmente marcadores de tiroiditis más que indicadores directos de función tiroidea. Actualmente, estudios comparativos entre la gammagrafía tiroidea y la prueba de estimulación con TRH han demostrado una mayor precisión diagnóstica de la gammagrafía, por lo que esta continúa siendo considerada el método gold standard para la evaluación diagnóstica del hipotiroidismo (Figura 15). El diagnóstico definitivo de hipotiroidismo suele establecerse mediante la integración de múltiples hallazgos clínicos, laboratoriales y, cuando es necesario, pruebas dinámicas complementarias1-3,11. Las alteraciones bioquímicas sistémicas también pueden aportar información útil. Muchos perros hipotiroideos presentan hipercolesterolemia, hipertrigliceridemia, anemia leve no regenerativa y ocasionalmente elevaciones moderadas de enzimas hepáticas3,8,11. En pacientes con sospecha clínica fuerte y perfiles hormonales compatibles, puede instaurarse tratamiento con levotiroxina y monitorizar cuidadosamente la evolución clínica1,3,11. Cuando el diagnóstico es correcto, la respuesta terapéutica suele ser favorable. Frecuentemente la mejoría del estado mental y de la letargia aparece en pocas semanas, mientras que las alteraciones dermatológicas requieren períodos más prolongados para resolverse, muchas veces entre 2 y 4 meses dependiendo de la severidad del cuadro y del ciclo folicular del paciente1,3,8,11. Por el contrario, la ausencia de mejoría clínica significativa tras un tratamiento correctamente instaurado debe hacer reconsiderar el diagnóstico inicial y reevaluar la posibilidad de enfermedades extratiroideas o de síndrome del eutiroideo enfermo1-3,11. En conjunto, el diagnóstico del hipotiroidismo debe entenderse como una integración clínica completa y no como la simple interpretación aislada de valores hormonales. Precisamente, el reconocimiento del síndrome del eutiroideo enfermo ha demostrado que múltiples enfermedades sistémicas pueden alterar profundamente las hormonas tiroideas sin que exista una verdadera insuficiencia primaria de la glándula tiroides1-4,6,10,11.



Bibliografía
- Spence S. Canine hypothyroidism: avoiding over diagnosing the condition. In Pract. 2022;44(2):68-75. doi:10.1002/inpr.74.
- Tappin S. Canine hypothyroidism: making a definitive diagnosis. Vet Ireland J. 2014;4(9):483-488. Sin DOI (no indicado).
- Scott-Moncrieff JC. Testing for hypothyroidism in dogs. Vet Clin North Am Small Anim Pract. 2007;37(4):647-669. doi:10.1016/j.cvsm.2007.05.015.
- Fliers E, Boelen A. An update on non-thyroidal illness syndrome. J Endocrinol Invest. 2021;44(8):1597-1607. doi:10.1007/s40618-020-01482-4.
- Giunti M, Troia R, Battilani M, Giardino L, Dondi F, et al. Retrospective evaluation of circulating thyroid hormones in critically ill dogs with systemic inflammatory response syndrome. J Vet Sci. 2017;18(4):471-477. doi:10.4142/jvs.2017.18.4.471.
- Crawford-Jennings MI. Thyroid function tests during nonthyroidal illness syndrome and recovery in acutely ill dogs. J Vet Intern Med. 2024;38(1):111-122. doi:10.1111/jvim.16947.
- Bolton TA, Panciera DL. Influence of medications on thyroid function in dogs: An update. J Vet Intern Med.2023;37(5):1626-1640. doi:10.1111/jvim.16823.
- Jaiswal M, Shukla PC, Tiwari A, Gupta D, Singh B, Maravi P, et al. Recent approaches in diagnosis and management of canine hypothyroidism: A review. Pharma Innov J. 2018;7(1):90-94. Sin DOI (no indicado en el artículo).
- Halsall DJ, Oddy S. Clinical and laboratory aspects of 3,3′,5′-triiodothyronine (reverse T3). Ann Clin Biochem.2021;58(1):29-37. doi:10.1177/0004563220969150.
- De Luca R, Davis PJ, Lin HY, Gionfra F, Percario ZA, Affabris E, et al. Thyroid Hormones Interaction With Immune Response, Inflammation and Non-thyroidal Illness Syndrome. Front Cell Dev Biol. 2021;8:614030. doi:10.3389/fcell.2020.614030.
- Boretti FS. Canine hypothyroidism: diagnosis and treatment. In: ESVE Summer School of Veterinary Endocrinology; 24 Jun 2018–30 Jun 2018; Bologna, Italy. p. 1-5. (handout/conference material).
- Harper A, Blackwood L, Mason S. Investigation of thyroid function in dogs treated with the tyrosine kinase inhibitor toceranib. Vet Comp Oncol. 2020;18(3):433-437. doi:10.1111/vco.12538.
- Feng HL, Li Q, Cao WK, Yang JM. Changes in thyroid function in patients with liver failure and their clinical significance: A clinical study of non-thyroidal illness syndrome in patients with liver failure. Hepatobiliary Pancreat Dis Int. 2020;19(6):561-566. doi:10.1016/j.hbpd.2020.05.001.
- Wolff EDS, Bilbrough G, Moore G, Guptill L, Scott-Moncrieff JC. Comparison of 2 assays for measuring serum total thyroxine concentration in dogs and cats. J Vet Intern Med. 2020;34:607-615. doi:10.1111/jvim.15703.
- Bolton TA, et al. Free thyroxine measured by chemiluminescence immunoassay and equilibrium dialysis in dogs with nonthyroidal illness syndrome. J Vet Intern Med. (ver DOI). doi:10.1111/ jvim.12573.
- Bennaim M, Shiel RE, Evans H, Mooney CT. Free thyroxine measurement by analogue immunoassay and equilibrium dialysis in dogs with non-thyroidal illness. Res Vet Sci. 2022;147:37-43. doi:10.1016/j.rvsc.2022.03.016.
- Daminet S, et al. Effect of an anti-inflammatory dose of prednisone on thyroid hormone monitoring in hypothyroid dogs. Vet Dermatol. 2011;22(3):246-251. doi:10.1111/ j.1365-3164.2010.00932.x.
- van Zyl E, et al. Serial changes in the concentrations of cortisol and thyroid hormones in Beagle dogs infected with Babesia rossi. Ticks Tick Borne Dis. 2023;14(2):102107. doi:10.1016/ j.ttbdis.2022.102107.